Métabolismes des microalgues
Il est maintenant important d'explorer le métabolisme algal car après ce qui vient d'être présenté, il est important de comprendre ce qui se passe à l'échelle des cellules dans le but d'améliorer ces "systèmes" de production de molécules d'intérêt, les optimiser et trouver des technologies innovantes qui leur soient adaptées.
L'image ci-dessous présente une coupe fine d'une cellule de micro-algue, et on distingue facilement le chloroplaste, où se déroule la photosynthèse.
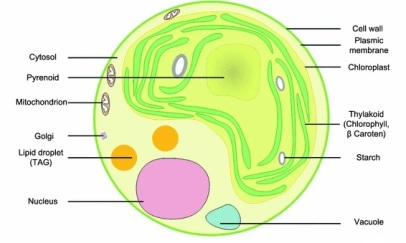
C : cytoplasme ; F : matière grasse ("fat") ; G : appareil de Golgi ; K : chloroplaste ; M : mitochondrie ; N : noyau.
La photosynthèse est un mécanisme complexe qu'il ne s'agit pas ici d'étudier de manière approfondie, mais d'en comprendre les principales étapes, sur la base du schéma ci-dessous. On y trouve deux grands groupes de réactions : les réactions de la phase lumineuse (réaction photochimique) et les réactions de la phase sombre (cycle de Calvin).
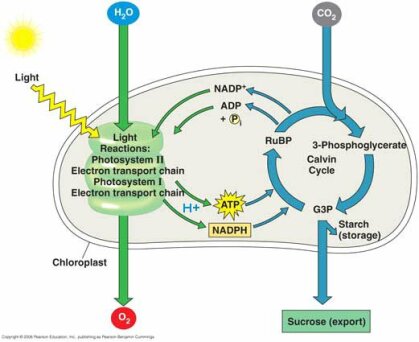
En captant l'énergie lumineuse, les micro-algues produisent de l'oxygène, du pouvoir réducteur et de l'ATP qui seront utilisés pour la fixation du CO2 au niveau du cycle de Calvin.
La réaction photochimique
Le schéma ci-dessous représente un chloroplaste et ses différentes parties.
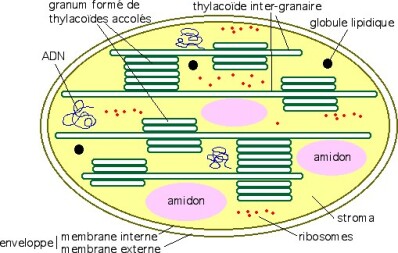
Tout se passe au niveau des membranes tylacoïdes (schématisées ci-dessous) où se situe le photosystème. Il s'agit de l'appareil photosynthétique composé de pigments, de protéines.
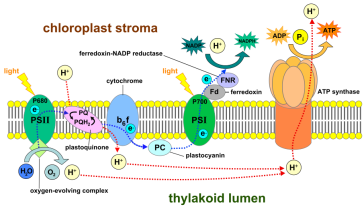
Le photosystème II représenté ci-dessous est composé d'une antenna contenant des pigments tels que la chlorophylle. Il contient également un complexe d'oxydation de l'eau ainsi qu'un complexe réactionnel. On trouve également une ATPsynthase indispensable à la synthèse de l'eau dans la cellule.
Parmi les pigments présents, on peut distinguer :
- la chlorophylle a, pigment majeure car c'est ce pigment qui va capter l'énergie lumineuse ;
- la chlorophylle b,
- les caroténoïdes,
- la phycobiliprotéine,
etc.
En observant les spectres d'absorption de ces pigments, on comprend que les micro-algues ne peuvent capter qu'une très faible fraction (400-700 nm) du spectre solaire.
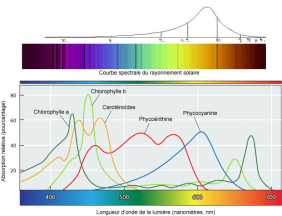
Photosystème II : La chlorophylle a, sous l'action des rayonnements lumineux (pic d'absorption à 680 nm), passe d'un état fondamental à un état excité, avec l'expulsion d'un électron qui sera transmis par une chaîne de transport d'électrons. Le schéma suivant représente cette chaîne de réactions.
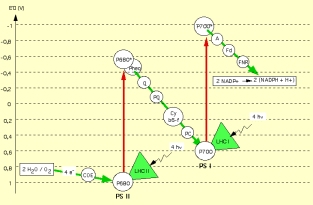
C'est une chaîne de réactions d'oxydo-réduction qui aboutit à la synthèse d'ATP.
Photosystème I : Le même mécanisme permet d'obtenir du pouvoir réducteur, sous forme de NADPH+H+ (ici le pic d'absorption des rayons lumineux se situe à 700 nm).
Une réaction d'oxydation de l'eau permet par ailleurs de fournir des électrons aux système.
Le cycle de Calvin
Les étapes du cycle de Calvin sont représentées ci-dessous :
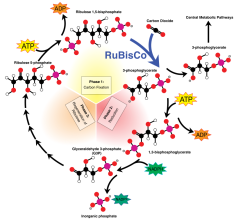
La fixation du CO2 s'effectue grâce à l'enzyme RuBisCO. Ensuite, on a une consommation de l'ATP et du pouvoir réducteur pour la catalyse de réactions de réduction permettant de générer le glycéraldéhyde 3-phosphate. Une partie de ce G3P sera recyclée dans le cycle de Calvin tandis qu'une autre sera exportée dans le cytoplasme, ce qui permet à la cellule de produire diverses molécules : glucides, acides aminés, etc.
Attention :
Le cycle de Calvin représente l'étape limitante dans le processus de photosynthèse. Il y a en effet des échelles de temps très différentes : la capture du CO2 est très rapide (pico ou nanoseconde), en revanche le transfert d'électrons est beaucoup plus lent (micro à milliseconde). En revanche, l'export vers le cytoplasme se situe à l'échelle de la seconde.
Calcul du rendement photosynthétique
On sait que 8 moles de photons sont nécessaires pour fixer une mole de CO2.
On a également la stoechiométrie globale de la synthèse de carbohydrates :
2NADPH + 3ATP + CO2 \(\rightarrow\) Carbohydrate
On sait par ailleurs que l'énergie moyenne d'une mole de photons est de 217 kJ.
Une mole de CO2 fixé représente environ 475 kJ
\(\Rightarrow\) L'efficacité théorique maximale de conversion de l'énergie lumineuse en énergie chimique dans la biomasse est donc de :\(\frac{475 kJ}{8\times 217 kJ}=27\)%
Attention :
Il ne faut pas oublier que seulement 45% de la lumière peut être exploitée par les micro-algues, ce qui ramène le rendement à environ 12% seulement. On verra plus loin que les rendements réels sont encore bien plus faibles que cette valeur.
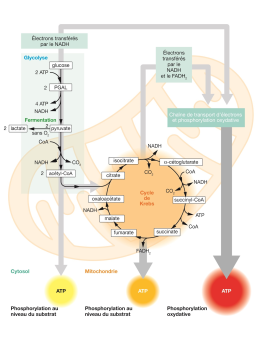
La respiration
Le mécanisme respiratoire a lieu au niveau des mitochondries, où le carbone intracellulaire sera consommé. Ce mécanisme prend une ampleur importance dans le cellule notamment en absence de lumière.
Effet des facteurs d'environnement sur les performances des procédés
Effet de la lumière
Bien entendu, les métabolismes sont ici présentés de manière très synthétique et n'abordent pas toute la complexité des réactions biochimiques mises en jeu, bien au delà de la photosynthèse et de la respiration. Ces réactions sont influencées par de nombreux paramètres tels que la concentration et la nature des nutriments, la température, l'intensité et la qualité de la lumière. Nous allons voir dans le suite que ce paramètres impactent à la fois les cinétiques de croissance et la composition des cellules.
On voit par exemple dans la figure ci-dessous le taux de photosynthèse en fonction de l'intensité de la lumière :
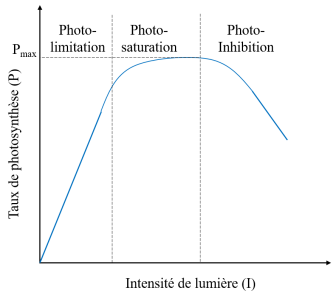
On observe trois grandes phases :
- une phase de photo-limitation : l'augmentation de l'intensité de la lumière entraîne une augmentation du taux de photosynthèse ;
- une phase de photo-saturation : la cellule est à sa capacité photosynthétique maximale ; l'augmentation de l'intensité de lumière n'entraîne pas d'augmentation de la photosynthèse ;
- une phase de photo-inhibition : les cellules peuvent même être endommagées par un excès d'énergie lumineuse.
\(\Rightarrow\) pour les deux dernières phases, on a une diminution importante de l'efficacité photosynthétique de l'algue.
En revanche, en présence d'un excès lumineux, certaines micro-algues réagissent à ce facteur de stress en produisant des molécules d'intérêt.
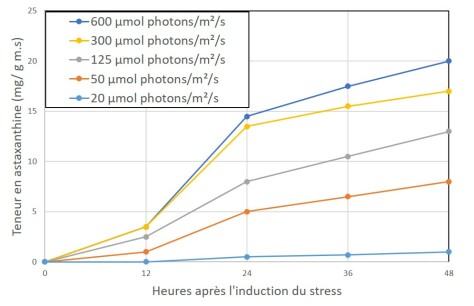
Il est donc important de comprendre les différentes voies métaboliques pour orienter et maîtriser la production industrielle de molécules d'intérêt. C'est le cas par exemple pour Haematococcus qui est verte à l'état végétatif et produit l'astaxanthine (rouge) lorsqu'elle est en présence d'un excès lumineux. On voit sur la figure ci-dessous que l'augmentation de l'intensité lumineuse pendant la culture entraîne une augmentation de l'accumulation d'astaxanthine.
Souche d'Haematococcus pluvialis soumise à différentes densités de flux de photons (courbes de bas en haut : 20, 50, 125, 300, 600 µmol photons.m-2.s-1)
On peut également observer un phénomène de photo-acclimatation qui consiste pour les cellules à adapter leur contenu en chlorophylle en réaction à la modification de l'intensité de lumière : face à une faible luminosité, les cellules vont augmenter leur teneur en chlorophylle (pour maintenir un niveau de photosynthèse suffisant).
Effet des nutriments - cas de l'azote
Dans le cas de la production de biocarburants par exemple, le procédé se déroule en deux phases principales : une phase de croissance puis une phase de production des molécules d'intérêt. Dans la première phase, le milieu de culture contient de l'azote, indispensable à la croissance. Dans la seconde phase, le milieu est dépourvu d'azote, et par réaction à ce déficit de nutriments, les cellules vont accumuler dans le cytoplasme des gouttelettes de lipides qui intéressent la production de biocarburants.